En Ikke-Tilfeldig Evolusjons-Hypotese (EITEH)-del II
(oversatt fra 'The Evolution Revolution' Kap. 2-slutten -av Dr. Lee M. Spetner, Jerusalem, Israel, Juni 2014.)
Bilde 1.  En Ikke-Tilfeldig Evolusjons-Hypotese
En Ikke-Tilfeldig Evolusjons-Hypotese
'Bevart DNA'
DNA kalles for 'bevart', om det (-ut fra det evolusjonære dogmet), er de samme sekvensene som er til stede i ulike arter. Da ser det enten ut som 'felles avstamning' eller 'gjenbruk av fungerende design'. Konklusjonen blir uansett at disse strengene må ha tjent en nødvendig funksjon, siden de er bevart og har nesten identiske DNA-strenger. Om disse felles DNA-strengene er gener, så koder de klart for protein eller DNA. Men, interessant nok er de best 'bevarte' strengene, ikke gener. De har fått navn av 'bevarte ikke-gen sekvenser' (Conserved NonGenic sequences -CNG). (Demitzakis et al. 2005) En sekvens er betraktet å være CNG, om den matcher minst 70% i to arter for minst 100 base-par (stigtrinn). Mellom 1 og 2% av genomet kvalifiserer som CNG, mens 'bevarte gener' okkuperer ca. 1%.
Bilde 2. Sjimpanse-menneske kromosomer ca 88% likhet
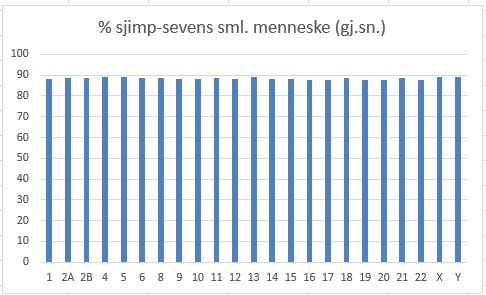 Det finnes også noen få sekvenser i genomet som har en enda høyere match arter i mellom, enn CNGs og kalles ultra-bevarte. Disse defineres som sekvenser med minst 200 base-bar (stigetrinn i DNAet) som er 100% identiske mellom f.eks. mus og mann.(Bejerano et al 2004). De må tjene en viktig funksjon, og siden det ikke er gener -så hva kan det være?
Det finnes også noen få sekvenser i genomet som har en enda høyere match arter i mellom, enn CNGs og kalles ultra-bevarte. Disse defineres som sekvenser med minst 200 base-bar (stigetrinn i DNAet) som er 100% identiske mellom f.eks. mus og mann.(Bejerano et al 2004). De må tjene en viktig funksjon, og siden det ikke er gener -så hva kan det være?
Nadav Ahituv og kolleger ved Lawrence Berkely National Laboratory i California, gjorde et eksperiment for å oppdage funksjoner ved slike sekvenser. De slettet fire slike sekvenser fra stamceller i muse-embryoet (Ahituv et al, 2007). Til sin overraskelse fant de ingen effekt på musene som ultra-bevarte strenger var fjernet fra -under ordinære omstendigheter. Det er imidlertid eksperimentelt bevis for at det finnes DNA-strenger som først spiller en rolle under ekstraordinære omstendigheter.
Et eksperiment ble utført med gjærceller, der hver av deres 6000 gener, ble slettet én etter én (Hillenmeyer et al, 2008). Av disse 6000 gener, ble 34% funnet å være nødvendige under normale omstendigheter. De gjenværende 66% viste ingen effekt under normale forhold. Men nesten alle (63% av totalen) viste vekstproblemer under omgivelses-endringer. De resterende 3% viste ingen effekt i dette eksperimentet, men kunne selvsagt gjort det i andre forhold i omgivelsene. Dermed er det sannsynlig at ca. 2/3 av de studerte gener inneholder gjærcellers innebygde evne å tilpasse seg omgivelses-endringer.
Ikke-tilfeldig evolusjon i celler
Ikke-tilfeldig evolusjon, som i EITHE, er blitt påvist å inntreffe i stor utstrekning i enkelt-cellede organismer, og spesielt i bakterier. Noen typer av endringer i bakterier er rapportert å inntreffe raskt, alt for raskt til å skyldes tilfeldige kopieringsfeil i genomet (punktmutasjoner). Men de kan forklares ut fra ikke-tilfeldige genetiske rearrangement, trigget ut fra endringer i omgivelsene.
Kryptiske gener
Kryptiske gener er gener som vanligvis er inaktive under et individs livssyklus, men kan aktiveres ved en epigenetisk endring (Hall et al. 1983). Et kryptisk gen har alle karakteristika til et ordinært gen, bortsett fra at de deaktiverte. Det skjer ved et DNA-segment som kalles en 'stilner' (silencer) (Schnetz 1995). Et slikt kryptisk gen har sin egen igangsetter (inducer) og stilner (repressor). Stilneren kan enten skrus av ved innsetting av et DNA-segment (Reynolds et al. 1986) eller ved sletting av et DNA-element (Parker et al. 1988, Schnetz 1995). Stilneren er under cellular kontroll, og påvirkes av omgivelsene.
Kryptiske gener er funnet i de fleste bakterie-typer og kan også bli funnet i høyere organismer. Minst 90% av E-coli bakterien har kryptiske beta-glukosid sukker-gener (Hall og Betts 1987, Hall 1999) Det er rapportert kryptiske gener som koder for enzymet acetohydroxyacid synthetase, som virker som katalysator i 1. trinn av dannelsen av aminosyrene valine, leucin og soleucin. (Mukerji og Mahadevan 1997). Varme-sjokk proteinet (Hsp90 er funnet å forårsake uttrykk av et kryptisk gen i bananfluen Drosophila, når den stimuleres av økt temperatur i omgivelsene (Rutherford og Lindquist 1998).
Kryptiske gener er også kjent å kode for antibiotika-resistans i bakterier (Hall, 2004). Disse kryptiske gener finnes vanligvis i plasmider i bakterien, og er inaktivt når ikke antibiotika er tilstede. I nærvær av antibiotika kan det inntreffe en epigenetisk endring, som tillater det kryptiske motstandsgenet å uttrykkes. For å unngå dette, må en prøve å finne ut hva i antibiotika som framkaller dette, og designe antibiotika rundt dette. Omgivelsene lokker fram innsettinger eller sletting av DNA-strenger, som gjør at det kryptiske genet kan uttrykkes. Endringene iverksettes altså når organismen blir påført endringer i omgivelsene (Hall 1999). Aktivering av kryptiske gener er eksempel på genetisk reorganisering, som svarer på input fra omgivelsene for å gjøre organismen mer tilpasningsdyktig for nye omgivelser.
 Nylon bakterien
Nylon bakterien
Bilde 3. Nyl-bakterier
Nylon ble oppfunnet i et forskningslaboratorium (DuPont) i Wilmington, Delaware 1935. Det begynte å produseres i 1939 for å bli brukt i damestrømper, samt for fiber, rep, nett og fallskjermer. I 1975 ble en bakterie (Flavobacterium) oppdaget i en dam avfallsvann til en japansk nylonfabrikk. Bakterien levde på avfall fra nylon, som da var ny i jordas bisofære (Kinoshita et al. 1975). I Japan hadde nylonproduksjon startet i 1951, så snaut 25 år tok det bakterien å tilpasse seg stoffet.
Bakteriene ble funnet å ha 3 enzymer, som sammen var i stand til å benytte nylonavfall i stoffskiftet (metabolization). Før nylon ble oppfunnet, hadde ikke Flavobacterium noe behov for disse enzymene. Enzymene syntes å ha dukket opp som tilsvart til knapphet på deres normal næring i nærvær av en overflod av nylon-avfall. To enzymer (E1 og E2) hydrolyserer en syklisk molekyl-forbindelse først til en lineær (E1) og så til 2 atskilte monomerer (E2). Genene som koder for disse to enzymene er kalt h.h.vis nylA og nylB (Kakudo 1993).
Oppdagelsen av disse genene i Flavolbatkerium-som oppsto på mindre enn 25 år, er tydelig ikke resultat av tilfeldige DNA-kopieringsfeil. I tillegg til den korte tilgjengelige tiden, måtte nylA og nylB begge måtte dannes, for at bakterien kunne benytte molekylforbindelsene i stoffskifte-sammenheng. Tilsynekomsten av de to genene syntes heller å være resultat av innebygd evne i bakterien til å respondere på input fra omgivelsene (EITHE). Dette er også bekreftet i et kontrollforsøk, foretatt langt utenom nevnte miljø. Enzymene som kunne benytte nylonavfallet i stoffskiftet, ble antagelig trigget av en epigenetisk begivenhet, som selv ble trigget av konsekvenser av mangel på næring.
Ikke-tilfeldige begivenheter i høyere organismer
Betingelser som forårsaker plager og lidelse i høyere organismer, kommer vanligvis fra mer komplekse forhold enn de som angår mikroorganismer. Effektene av slike betingelser, blir vanligvis formidlet via hjernen. Eks. på miljømessig stress og forstyrrelser, omhandler rovdyr som blir for dominerende og alvorlig klimatiske endringer. Det er kjent at i fisk, så forårsaker stress endringer i både oppførsel og fysiologi (Iwama, 1998). Disse endringene tjener vanligvis til å redusere stress for fisken. Komplekse miljømessige endringer kan trigge epigenetiske endringer, i tråd med en Ikke-Tilfeldig Evolusjonær mekanisme. I følge Iwama foregår stress-respons i fisk i tre etapper: i) Frigjøring av spesielle stress-hormoner (catecholaminer) i blodet. Disse fungerer som første-budbringere. ii) Så ferdes disse hormonene rundt i blodet til alle kroppens celler (andre-budbringere). iii) Så trigger disse andre-budbringerne passende enzym-aktivitet (Cohen 1988), som kan påvirke både fysiologi og oppførsel for å redusere virkninger av omgivelses-stress. Andre-budbringerne kan slå celler av og på. Genetiske endringer kan observeres på inidivid-nivå, som vekst eller reproduksjon, eller i hele populasjonen.
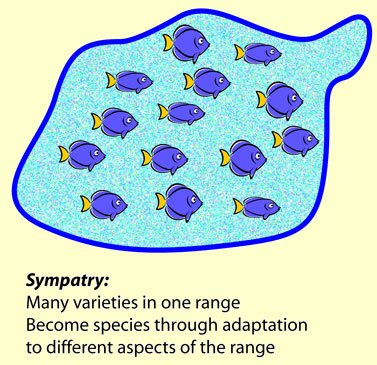 En slik Ikke-Tilfeldig Evolusjonær mekanisme kan resultere i nylig dannede arter (species) innen samme livsområde som originale arter (sympatric speciation). Mange eks. på rask utvikling har vist seg å være eks. på 'sympatric speciation', med oppdukking av en isolerende mekanisme som forhindrer nye arter fra å bli fortynnet av eksisterende arter. På slutten av 1900-tallet hadde det samlet seg nok eks. på slike nylig dannede arter, til at selv motstandere (Mayr 2004) måtte vedgå eksistensen og viktigheten av det. Neo-darwinister ble tvunget til å finne modeller for hvordan dette kunne skje. Det ble foreslått f.eks. immigrasjon inn i en eksisterende bestand. Det kunne selvsagt inntreffe, men kunne ikke alene gjøre rede for de fleste eks. av nylig dannede arter (sympatric speciation). Tilfeller av en slik rask artsdannelse kan forklares ut fra omgivelses-induserte, epigenetiske begivenheter av en Ikke-Tilfeldig Evolusjonær mekanisme , men ikke ved tilfeldige mutasjoner og naturlig seleksjon.
En slik Ikke-Tilfeldig Evolusjonær mekanisme kan resultere i nylig dannede arter (species) innen samme livsområde som originale arter (sympatric speciation). Mange eks. på rask utvikling har vist seg å være eks. på 'sympatric speciation', med oppdukking av en isolerende mekanisme som forhindrer nye arter fra å bli fortynnet av eksisterende arter. På slutten av 1900-tallet hadde det samlet seg nok eks. på slike nylig dannede arter, til at selv motstandere (Mayr 2004) måtte vedgå eksistensen og viktigheten av det. Neo-darwinister ble tvunget til å finne modeller for hvordan dette kunne skje. Det ble foreslått f.eks. immigrasjon inn i en eksisterende bestand. Det kunne selvsagt inntreffe, men kunne ikke alene gjøre rede for de fleste eks. av nylig dannede arter (sympatric speciation). Tilfeller av en slik rask artsdannelse kan forklares ut fra omgivelses-induserte, epigenetiske begivenheter av en Ikke-Tilfeldig Evolusjonær mekanisme , men ikke ved tilfeldige mutasjoner og naturlig seleksjon.
Bilde 4. Tilpasning kan frambringe fruktbarhets-sperrer
I århundrer har naturalister observert at dyrepopulasjoner og planter endrer seg med omgivelsene. Foreslåtte forklaringer på disse fenomen har vanligvis inkludert arv av oppnådde egenskaper (Waddington 1942), hvorav Lamarcks forslag er det mest kjente. J.M. Baldvin foreslo i 1896 en forklaring hvorav tilpasset oppførsel oppsto fra miljømessige endringer. Det gikk i korthet ut på at evnen til å lære og videreføre kunnskap var til stede. En Ikke-TIlfeldig Evolusjonær mekanisme (EITEM) kan redegjøre for arvelige endringer ved at miljømessige endringer (stress) framkaller hormonproduksjon som når fram til alle celler, også kjønnsceller, og fremkaller et tilpasset svar på miljøendringene. Denne beskrivelsen er basert på raskt forekommende, observerte fenomen i populasjoner, som kan forklares mye bedre slik (EITEM), enn ved tilfeldige mutasjoner i enkeltindivider som ville kreve lang tilpasningstid i populasjonen som helhet.
Evnen ved en organisme til å endre sitt utseende og oppførsel (inkl. i fenotype-begrepet), kalles ofte for fenotype-plastisitet. I det ligger at når de miljømessige endringer er tilbakestilt, så vil fenotypen returnere til sin opprinnelige tilstand (sml. Darwins finker). Imidlertid kan også disse fenotype-endringene også være arvelige, som indikerer at de har en (epi-)genetisk basis. I senere år er det kommet en del dokumentasjon på hvordan miljømessige endringer kan resultere i nye, arvelige egenskaper:
 a) Omgivelsesendringer kan forårsake endrings-stress i en organisme. b) Stress kan forårsake genetiske rearrangeringer, som kan være tilpassede. Det kan foregå på flg. måter: i) Frigjøring av spesielle stress-hormoner (catecholaminer) i blodet. Disse fungerer som første-budbringere. ii) Så ferdes disse hormonene rundt i blodet til alle kroppens celler -andre-budbringere. iii) Dernest trigger disse andre-budbringerne passende enzym-aktivitet (Cohen 1988), som kan påvirke både fysiologi og oppførsel for å redusere virkninger av omgivelses-stress. Andre-budbringerne kan slå celler av og på. Genetiske endringer kan observeres på inidivid-nivå, som vekst eller reproduksjon, eller i hele populasjonen.
a) Omgivelsesendringer kan forårsake endrings-stress i en organisme. b) Stress kan forårsake genetiske rearrangeringer, som kan være tilpassede. Det kan foregå på flg. måter: i) Frigjøring av spesielle stress-hormoner (catecholaminer) i blodet. Disse fungerer som første-budbringere. ii) Så ferdes disse hormonene rundt i blodet til alle kroppens celler -andre-budbringere. iii) Dernest trigger disse andre-budbringerne passende enzym-aktivitet (Cohen 1988), som kan påvirke både fysiologi og oppførsel for å redusere virkninger av omgivelses-stress. Andre-budbringerne kan slå celler av og på. Genetiske endringer kan observeres på inidivid-nivå, som vekst eller reproduksjon, eller i hele populasjonen.
Bilde 5. Pupfish i Death Valley reservat
Som eks. kan nevnes at for ett bestemt fiskeslag (pupfish), så påvirket selv små forskjeller i oppveksttemperatur utvikling av og kroppsbygning til pupfish. Endringer i omgivelser påvirket produksjon av hormonet thyroid, som igjen påvirket kroppsbygning. Når omgivelser ble endret, påvirket det produksjonen av et annet hormon (arginine vasotoin -AVT) som spilte en viktig rolle i fiskens oppførsel. Cellene som produserer AVT befinner seg i hjernen, og er kjent for å respondere på stress. Disse observerte endringene skjedde ved flytting av fisk, og frambringer støtte for en rask, Ikke-TIlfeldig, Evolusjonær mekanisme (EITEM). Denne mekanismen forklarer endringene som respons av en inne-bygd egenskap ved organismen, for å tilpasse den til langsiktige endringer i omgivelsene. Hvordan en slik mekanisme oppsto vet vi for tiden ikke, og det må utsettes til videre studier.
Oversatt og tilpasset .htm-format ved Asbjørn E. Lund